PRESS RELEASES

『ガラクタ』RNAの遺伝子活性化における新しい役割研究成果

『ガラクタ』RNAの遺伝子活性化における新しい役割
|
1.タイトル:「ガラクタ」RNAの遺伝子活性化における新しい役割
2.発表概要:
ゲノム情報の「暗黒物質(ダークマター)」といわれる非翻訳型RNA(ノンコーディングRNA)の出現とともに、段階的にクロマチン構造が緩み、遺伝子の発現が活性化される機構が、国立大学法人東京大学(小宮山宏総長)と独立行政法人理化学研究所(野依良治理事長)の共同研究により、世界で初めて明らかにされた。
3.発表内容:
発見の背景
ヒトゲノム解析結果から、下等な生物と比較して、ヒトの遺伝子数がそれほど多くないことが示された。これは、生物の複雑化や多様化において、遺伝子数の増大よりも、遺伝子発現制御の複雑化が重要であることを示している。また、昨今の網羅的な遺伝子発現の解析から、タンパク質をコードしていない転写物(非翻訳型RNAまたはノンコーディングRNA)が、予想以上に多く存在することが示された。非翻訳型RNAは、当初何をしているか不明であったため「ガラクタ」のような存在ではないかと考えられていた。しかし、最近になって、発生・分化に応じて転写制御を受けるほか、遺伝子発現の抑制などの機能が示されつつあり、さまざまな遺伝子制御過程に重要な役割を持つと考えられるようになった。このような無数の非翻訳型RNAの発見は、生命科学に大きな転換点をもたらしつつあり、「RNA新大陸発見」と称されるに至っている。しかしながら、非翻訳型RNAの遺伝子発現制御における役割は不明な点が多く、特にmRNA型の非翻訳型RNAの機能については、ほとんど明らかになっていない。
太田邦史(東京大学大学院総合文化研究科教授/理化学研究所客員主幹研究員)の研究室では、酵母を用いてDNA組換えとクロマチン構造※1の関係を調べてきた。その過程で、DNA組換えが頻発する場所で、減数分裂時にクロマチン再編成が起こることを示した。また、このクロマチン再編成には、ある種の配列特異的DNA結合タンパク質(CREB/ATF型転写因子※2)が関わることを見出した。さらに、太田邦史教授と廣田耕志氏(元 理化学研究所 基礎科学特別研究員)は、同様なクロマチン再編成の仕組みが、グルコースが枯渇した環境下(グルコース飢餓)にある分裂酵母のfbp1遺伝子(フルクトース-1,6-ビス脱リン酸酵素※3)の転写プロモーター領域にも認められ、fbp1遺伝子の活性化に関わることを明らかにしていた。
発見の詳細
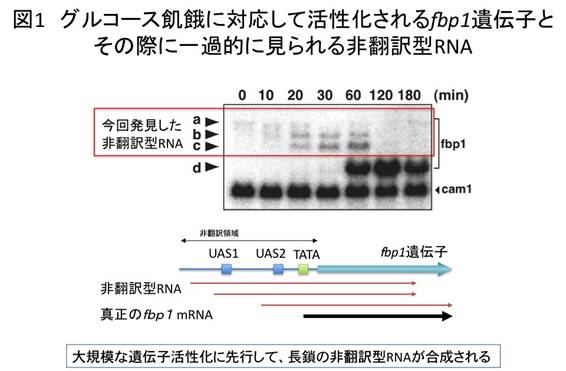
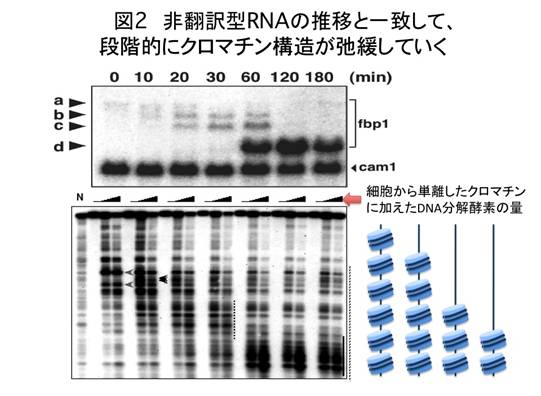
fbp1遺伝子は、グルコースが培地に存在する間はほとんど転写されない。ところが、グルコースが培地から失われると(グルコース飢餓)、1時間ほどで顕著に活性化される。今回廣田耕志氏と太田邦史教授らは、グルコース飢餓状態に移行する際、微量の非翻訳型RNAがあらかじめ転写されていることを発見した(図1)。この非翻訳型RNAは、正規のfbp1遺伝子プロモーターのさらに上流域から転写される長鎖のmRNA型非翻訳型RNAであり、タンパク質には全く翻訳されない。興味深いことに、グルコース飢餓に対応してfbp1遺伝子の活性化がはじまると、転写開始部位がfbp1コード領域に近接したいくつかの転写開始点に順次移行し、RNA量の増大に相反して、長さがだんだんと短くなっていった。これに呼応するように、fbp1プロモーター領域のクロマチン構造が徐々に開いた状態に移行していくことも示された(図2)。グルコース飢餓から1時間ほどすると、正規のfbp1転写開始点から大量のmRNAが合成され、タンパク質への翻訳が始まった。この時期におけるfbp1領域のクロマチン構造は、広い範囲でヌクレアーゼの消化を受けやすい開いた状態をとっていた。
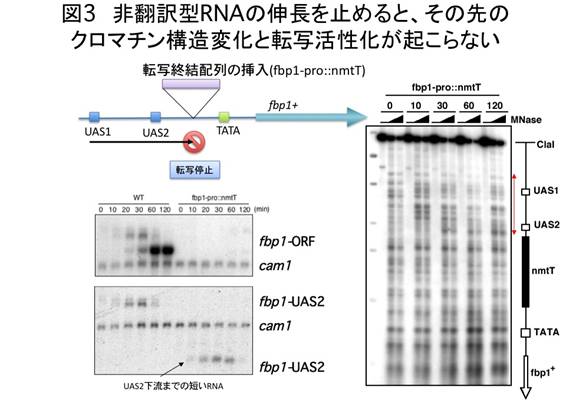
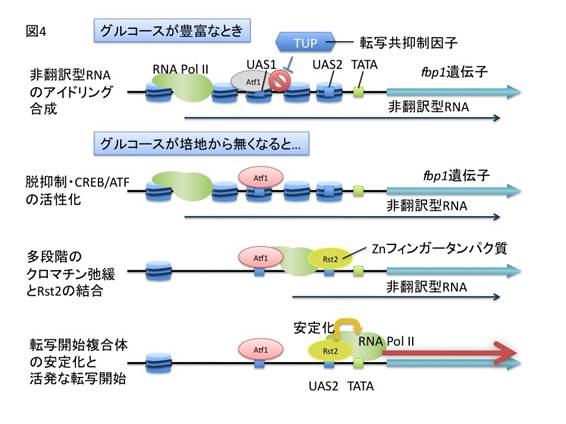
上記の結果は、長鎖非翻訳型RNAがRNAポリメラーゼII※4によって合成される過程で、順次fbp1プロモーター領域のクロマチン構造が弛緩していき、これがカスケード的に生じることで、転写が段階的に活性化される可能性を示唆している。そこで、この考えを検証するため、fbp1プロモーター領域の複数の箇所に転写終結配列※5を挿入した酵母株を作製し、fbp1の発現やクロマチン構造を解析した。その結果、長鎖非翻訳型RNAの合成を途中で終結させると、大規模なfbp1の転写活性化が起こらなくなること、また長鎖非翻訳型RNAの終結点以降でのクロマチン再編成が起こらなくなることを確認した(図3)。分子レベルの解析で、長鎖非翻訳型RNAの転写には、RNAポリメラーゼII、fbp1プロモーター領域に結合するCREB/ATF型転写因子、C2H2Znフィンガータンパク質※6、Groucho型の転写共抑制因子※7が協調的に関わることが示された(図4)。さらに、分裂酵母の全ゲノムをカバーするゲノムタイリングDNAチップ※8を用いて、グルコース飢餓で転写が誘導されるほかのいくつかの遺伝子でも、同様な長鎖非翻訳型RNAが誘導初期に転写されることを明らかにした。
以上の結果から、非翻訳型RNAの転写を伴う段階的なクロマチン再編成が、遺伝子の活性化にも重要な役割を果たすことが示された。
意義と波及効果
CREB/ATF型の転写因子や、Groucho型転写共抑制因子は、糖代謝のほか、高等真核生物では発生や分化にも関わることが知られている。また、記憶に必要な神経細胞の長期増強の際にも、CREB/ATF型の転写因子が関わるクロマチン再編成が起こることが報告されている。これらのことから、今回発見したタイプの非翻訳型RNAは、おそらく発生や分化、長期記憶などの過程に関わる遺伝子群においても活躍しているものと考えられる。近年のクロマチンをベースにした研究で、真核生物の転写制御機構の概念は、大腸菌をモデルとしたジャコブとモノーのオペロン説※9から大きな発展を見せつつある。今回発見された機構も、真核生物の遺伝子制御に関する研究に、新しい展開をもたらすものと期待される。また、ヒトなどで同様な機構を調べることで、糖尿病などの代謝異常疾患や、人間の記憶の仕組みが解き明かされる可能性がある。
4.発表雑誌:
9月28日付けのNature誌オンライン版で発表。
Hirota K., Miyoshi T., Kugou K., Hoffman C.S., Shibata T., and Ohta K.
Stepwise chromatin remodeling by a cascade of transcription initiation of non-coding RNAs Nature, in press (2008)
5.注意事項:
報道の解禁 日本時間9月29日 午前2時(新聞は9月29日朝刊)
6.問い合わせ先:
東京大学大学院総合文化研究科 広域科学専攻
教授 太田邦史
URL: http://bio.c.u-tokyo.ac.jp/labs/ohta/
7.用語解説:
※1クロマチン構造:真核生物のゲノムDNAは、ヒストンやそれ以外のタンパク質と結合し、高度に凝縮した状態で存在する。このような構造をクロマチン構造と呼ぶ。局所的なクロマチン構造の変化を介して、転写因子などのタンパク質の染色体DNAへの接近のしやすさが制御される。クロマチン構造は、転写や組換え・複製などの遺伝情報制御において、中心的役割を果たすことが示されつつある。
※2CREB/ATF型転写因子(CREB: CRE-binding protein、ATF:activating transcription factor):cAMP 応答配列(CRE: cAMP responsive element)に結合する、塩基性ロイシンジッパーを持った配列特異的DNA結合タンパク質である。同類の転写因子が酵母からヒトまで広く存在しており、糖代謝調節、ストレス応答や発生、神経の長期増強などに重要な役割を果たす。
※3フルクトース-1,6-ビス脱リン酸酵素:酵母をはじめ、多くの真核生物では、エネルギー源としてのグルコース(ブドウ糖)が枯渇すると、ほかの栄養素から糖新生というはたらきによってグルコースの供給を維持する。この際、グルコースを代謝する解糖系の逆反応を用い、その逆反応の1つに不可欠な酵素の1つが、フルクトース-1,6-ビス脱リン酸化酵素である。この酵素は、生育環境にグルコースが存在すると転写が低く抑えられ,ほとんど細胞内に存在しないが、グルコース飢餓に際して大量に合成されるようになっている。したがって、この過程には、厳密な遺伝子活性の制御が必要であり、遺伝子発現研究のための良いモデル系を提供する。
※4RNAポリメラーゼII(RNA Pol II):真核生物のRNA合成を担当する酵素は3種類ある。そのうちの1つがRNA Pol IIであり、基本的にはタンパク質をコードする遺伝子領域(クラスII遺伝子)の転写を行う。それ自身では、転写開始点に特異的に結合できず、転写活性化因子を介して転写開始点のクロマチンに結合する。
※5転写終結配列:遺伝子の3´側に配置する配列で、転写されたRNAでは末端部分に位置し、RNAポリメラーゼの反応を終結させる機能がある。
※6C2H2Znフィンガータンパク質:亜鉛を配位することでDNA結合活性を示すドメインを持つタンパク質のうち、亜鉛の配位が2つのシステインと2つのヒスチジンによるもの。遺伝子発現に重要な役割を果たす。
※7Groucho型の転写共抑制因子:Groucho(グルーチョ)という転写共抑制因子は、高等真核生物のWnt シグナルの下流に位置する転写制御因子である。高等生物の体軸・体節形成に関与する。当初、ショウジョウバエの体節剛毛が太くなる変異の原因遺伝子(名は眉毛の濃い喜劇俳優から取られた)として同定された。酵母などでは、Tupといわれる転写共抑制因子が同類であり、ストレス応答遺伝子などの転写制御に関わる。
※8ゲノムタイリングDNAチップ:染色体の配列を、端から端までもれなく網羅して、20塩基対や100塩基対単位に区切り、その区画に対して25塩基程度のオリゴDNAを設計し、個々にチップ上に合成したもの。染色体のどの部分が転写されているかが、高解像度で網羅的に把握できる。
※9オペロン説:オペロン説は、大腸菌のラクトース・オペロンの研究から、ジャコブとモノーによって1961年に提唱された遺伝子制御理論。遺伝子の発現は、構造遺伝子の上流に位置する調節領域に転写調節タンパク質が結合・脱離することにより、転写レベルで制御される、という概念。
8.添付資料: