PRESS RELEASES

腸管における病原細菌(赤痢菌)と宿主自然免疫のあらたな攻防を解明:新規病原因子の立体構造とその標的宿主因子の発見研究成果

腸管における病原細菌(赤痢菌)と宿主自然免疫のあらたな攻防を解明: |
平成24年3月12日
東京大学医科学研究所
1. 発表者:
真田 貴人1、 Kim Minsoo1、 三室 仁美2、 鈴木 仁人3、 小川 道永3、 小山 晃穂3、 芦田 浩3、 小林 泰良3、小山 智洋4、 長井 信也4、 柴田 佑里5、 合田 仁5、 井上 純一郎5、 水島 恒裕6*、 笹川 千尋 1, 3, 4*
1. 東京大学医科学研究所 感染症国際研究センター 感染制御系
2. 東京大学医科学研究所 感染症国際研究センター?感染制御系 細菌学
3. 東京大学医科学研究所 感染・免疫部門 細菌感染分野
4. 財団法人日本生物科学研究所
5. 東京大学医科学研究所 癌・細胞増殖部門 分子発癌分野
6. 兵庫県立大学 生命理学研究科 生命科学専攻 生体物質構造解析学部門
* Correspondence authors: 笹川千尋(東大医科研)、水島常裕(兵庫県立大)
2.発表のポイント:
◆どのような成果を出したのか:
「腸管における病原細菌と宿主自然免疫のあらたな攻防を解明」
?赤痢菌が分泌するあらたな酵素とそれによる腸管感染早期の炎症制御の発見?
◆新規性(何が新しいのか)
赤痢菌(注1)の未知病原因子(新規酵素)の解明により、腸粘膜の病原体侵入に対する新規な生体防御応答と、これに対する病原細菌の対抗戦略を発見した。
◆ 社会的意義/将来の展望
本研究で明らかにされた炎症シグナルや病原因子を新たな標的として(あるいはその原理を踏まえて)、炎症性腸疾患や腸管感染症を早期に阻止する創薬や弱毒ワクチン開発に寄与する。
3.発表概要:
毎年約1500万人が感染症で命を失われ、そのなかの約200万人は腸管感染症が原因である。腸管は無数の微生物に常に暴露されているが、微生物の侵入から生体を守るために自然免疫を中心とする堅固な防御システムが幾重にも備えられている。一方、赤痢菌やその仲間である病原性大腸菌(O157等)等の腸管病原細菌は、それら防御システムを巧みに回避して生体へ侵入する高度に進化した機能を備えている。病原細菌の腸粘膜への侵入に対する自然免疫応答と、それを病原細菌がどのようにして回避するか、そのメカニズムは全く不明であった。
今回、東京大学医科学研究所の笹川千尋教授と真田真人研究員らは、兵庫県立大学の水島恒裕教授および大型放射光施設(SPring-8)との共同研究により、赤痢菌をモデルにして、(i) 粘膜上皮に対する病原体の侵入認識機構。(ii) この宿主認識・防御に対抗する赤痢菌の手段 を明らかにした。具体的には、(i) 細菌が細胞侵入するときに形成される葉状突起(ラッフル膜)を危険信号として認識する仕組みとして、「葉状突起に局在するジアシルグリセロール(注4)(DAG)-TRAF6-NF-κBに依存した炎症シグナル経路が重要である」ことを明らかにした。(ii) 赤痢菌の対抗手段として菌から分泌されるOspIを同定し、そのタンパク質の立体構造(図2参照)および生化学的性状を解明した。(i)と(ii)の結果から、OspIは、上述の炎症シグナル経路の制御に重要なTRAF6の活性化に必要なUBC13に結合性して、UBC13の100番目のグルタミンを脱アミド化する「新規な脱アミド化酵素である」ことを発見した(図1参照)。本研究により、感染初期の病原体に対する粘膜上皮(注2)の新規な防御機構と、それに対抗する病原体側のあらたな戦術が解明され、これを標的に創薬やワクチン開発への応用が期待される。
添付資料:
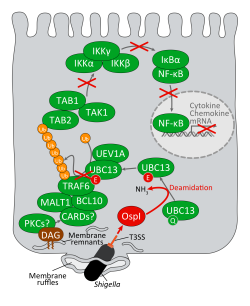
図1 赤痢菌のOspIによる炎症抑制メカニズム
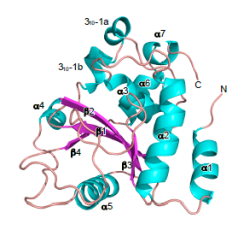
図2 OspIの立体構造
図の解説
図1 赤痢菌のOspIによる炎症抑制メカニズム
赤痢菌の上皮細胞侵入によりラッフル膜(葉状突起;マクロピノサイトーシス)が菌周囲に形成される。菌はラッフル膜を破壊して最終的に細胞質へ移行する。上皮細胞の側底面にラッフル膜が形成されるとジアシルグリセロールがラッフル膜に局在化する。ジアシルグリセロール(注4)は、プロティンキナーゼC (PKCs)によるシグナルのセカンドメッセンジャーとして働き、その結果CMB(CARD-MALT1-BCL10)複合体-TRAF6- NF-κB経路が活性化され、抗菌性ペプチド、炎症性サイトカインやケモカイン(注6)の産生が誘導される。赤痢菌はこれを回避して細胞質で生存・増殖を行うために、細胞侵入時にIII型分泌装置(注3)よりOspIエフェクター(病原因子)を分泌する。OspIは、UBC13に特異的に結合して、UBC13の100番目のグルタミンを脱アミド化してそれをグルタミン酸へと改変する。その結果、UBC13は失活する。UBC13はTRAF6の活性化因子であり、UBC13の活性化が失われるとTRAF6の活性化が阻止され、その下流のNF-κBシグナル経路も働かなくなる。このようにして、赤痢菌は感染の開始早々に、宿主の炎症・自然免疫応答(注4)を抑制する。その結果、赤痢菌の粘膜上皮内への感染が促進する。
図2 OspIの立体構造
赤痢菌からIII分泌装置を介して分泌されるOspIタンパク質の結晶化に成功した。その結晶構造解析をSPring-8で行い、2.0 A分解能でのOspI立体構造解析に成功した(Protein Data BankにID番号: 3B21で登録済み)。OspIは、4個のβ-ストランド (赤)、7個のα-ヘリックス (青)、1個の310 ヘリックス (青)からなる構造をもつ事が明らかとなった。OspIの立体構造を、既に報告されている様々なタンパク質の一次配列と立体構造と比較した。その結果、OspIのアミノ酸の一次配列と類似するタンパク質は見いだせなかったが、OspIの立体構造は、シュードモナス‐シリンゲ(赤痢菌と同様にIII分泌装置をもつ植物病原細菌)の病原因子として知られるAvrPphBと類似性が認められた。AvrPphBタンパクも、OspIと同様にシステイン(Cys)、ヒスチジン(His)、アスパラギン酸(Asp)からなる活性中心をもつプロテアーゼ酵素である。これを手掛かりに、OspIの示すUBC13に対する脱アミド化活性に関わるCys-His-Aspの3アミノ酸残基の重要性を、各アミノ酸置換変異体を作製して明らかにした。
4.発表内容:
研究の背景・先行研究における問題点
腸管における微生物と宿主の攻防の中で自然免疫(注5)は生体防御の中心的な役割を担っている。病原体は、腸管へ侵入後、粘膜上皮で増殖して定着するまでの総ての過程において自然免疫により監視され、その結果、炎症性サイトカイン、ケモカイン(注6)の発現が誘導され、病原体が無防備であれば排除されてしまう。したがって、感染初期の自然免疫応答は生体防御に極めて重要であるが、これまでは主にマクロファージを中心とする免疫応答の研究が主体であった。また、細菌の上皮細胞への感染における自然免疫応答およびこれに対抗する病原細菌の戦術に関して、多くが感染の後期における情報が蓄積されてきたが、感染初期の免疫応答に関する情報は乏しかった。一方、多くの病原細菌は粘膜上皮細胞を足場として感染し下痢をはじめとするさまざまな炎症性疾患を引き起こす。したがって病原細菌は様々な戦略によって炎症・自然免疫応答を適切に制御することが感染・定着に必須であることが知られてきた。しかし感染開始時の腸粘膜における自然免疫応答とそれに対抗する病原細菌の戦術に関する知見は皆無であった。これまでも赤痢菌をはじめとする腸管病原細菌は、III型分泌装置(注3)と呼ばれるタンパク分泌装置によって、エフェクターと称する一群の病原因子を宿主細胞に分泌し、抗菌性ペプチド、炎症性サイトカイン、ケモカインの発現を抑制することが知られていた。今回我々は、赤痢菌のエフェクターの中に、菌が宿主侵入時に炎症応答を阻害するエフェクターを分泌していることを見いだし、その立体構造と生化学的性状の解析を手掛かりの以下の発見が導きだされた。
研究内容
1. プロテオミクス解析(注7)によって、赤痢菌のIII 型分泌装置から分泌される新規タンパク質の探索を行った結果、菌のIII型分泌装置によって分泌されるタンパク質のなかに未同定のOspIが存在することを見いだした。
2. マイクロアレイ解析(注8)を用いて、赤痢菌の上皮感染におけるOspIの役割について検討した。野生型およびospI欠損赤痢菌を感染させた上皮細胞について、遺伝子の発現量の変化をマイクロアレイ解析によって網羅的に比較した。その結果、ospI欠損株感染細胞において一連のケモカイン (IL-8など) と炎症性サイトカイン (TNFαなど) の顕著な発現量増加が認められた。即ち、OspIは「炎症抑制作用」を持つことが示唆された。
3. NF-κBの活性化経路は宿主の初期免疫応答すなわち、炎症性サイトカイン・ケモカイン産生誘導に必須である。そこでNF-κB経路の活性化におけるOspIの作用について解析した。宿主細胞への赤痢菌侵入に伴い、侵入部位にジアシルグリセロール(DAG)の集積が認められる事を見いだし、DAGの集積が、その下流のDAG-CBM (CARDs-BCL10-MALT1) 複合体-TRAF6-NF-κB経路の活性化の引き金となる事を示した。さらに、OspIはDAG-CBM複合体-TRAF6- NF-κB経路を抑制する活性を有する事を示した。
4. OspIは、細胞内でTRAF6の自己ユビキチン化を抑制する事によってTRAF6の活性化を抑制する働きがあった。さらにOspIによるTRAF6活性化の抑制分子機構について検討した結果、OspIはTRAF6 (ユビキチン転移酵素) のE2 (ユビキチン結合酵素) であるUBC13を標的としていた。OspIは、UBC13の100番目のグルタミン残基を脱アミド化修飾してグルタミン酸に変換する脱アミド化酵素であることが明らかになった。事実、OspIによって脱アミド化修飾を受けたUbc13のE2活性は、顕著に低下している事を明らかにした。
5. 本研究と平行して、水島恒裕教授(兵庫県立大学)はOspIの結晶化に成功し、兵庫県のSPring-8(大型放射光施設)でOspIの結晶構造の解析を行った。その結果、OspIはシステインプロテアーゼ様の立体構造をもち、システイン (C)、ヒスチジン (H)、アスパラギン酸 (D) からなる活性中心 (C-H-D triad) を持つ新規なタンパク質であることが明らかとなった。
6. OspIの立体構造による知見に基づきC-H-D triadの点変異解析を行った。その結果、C-H-D triadの各点変異体は、OspIによるUBC13の脱アミド化に必須であることが示され、立体構造解析から予測された結果と完全に一致した。
社会的意義・今後の予定
本研究によって感染初期の病原体に対する粘膜上皮の新規な防御機構と、それに対抗する病原体側のあらたな戦術が解明された。得られた病原体と宿主側の知見は、炎症性腸疾患の新たな治療標法の開発に繋がることが期待される。また下痢関連疾患に関わる腸管感染症の病原細菌に対するワクチンの開発や発症予防法にも貢献すると思われる。
5.発表雑誌:
雑誌名:「Nature」(3月11日, Advance Online Publication 18:00 ロンドン現地時刻)
論文タイトル:
The Shigella flexneri effector OspI deamidates Ubc13 to dampen the inflammatory response
著者:
真田 貴人1、 Kim Minsoo1、 三室 仁美2、 鈴木 仁人3、 小川 道永3、 小山 晃穂3、 芦田 浩3、 小林 泰良3、小山 智洋4、 長井 信也4、 柴田 佑里5、 合田 仁5、 井上 純一郎5、 水島 恒裕6*、 笹川 千尋 1, 3, 4*
1. 東京大学医科学研究所 感染症国際研究センター 感染制御系
2. 東京大学医科学研究所 感染症国際研究センター?感染制御系 細菌学
3. 東京大学医科学研究所 感染・免疫部門 細菌感染分野
4. 財団法人日本生物科学研究所
5. 東京大学医科学研究所 癌・細胞増殖部門 分子発癌分野
6. 兵庫県立大学 生命理学研究科 生命科学専攻 生体物質構造解析学部門
* Correspondence authors: 笹川千尋(東大医科研)、水島常裕(兵庫県立大)
DOI番号:10.1038/nature10894
6.問い合わせ先:
笹川千尋 教授
〒108-8639 東京都港区白金台4-6-1
東京大学医科学研究所 感染・免疫部門 細菌感染分野
7.用語解説:
(注1) 赤痢菌:分類学的には大腸菌と同じ仲間。赤痢菌は細菌性赤痢の原因菌。昨年8月、本菌の感染によりチェーンレストランで赤痢が発生した。赤痢菌は腸管下部(大腸・直腸)の吸収上皮細胞へ侵入することにより感染を開始し、ついで上皮細胞内で増殖しつつ周囲の上皮細胞へ拡散するが、一方で感染は自然免疫により検知され腸管に炎症が起こる。
(注2) 粘膜上皮:腸管などの管腔状臓器の単層の上皮細胞。腸管の管腔側で栄養を吸収する役割と共に、微生物の侵入の対するバリアー(防御機構)として重要な役割も果たしている。
(注3) III型分泌装置:多くの病原細菌が保有する特殊な病原因子(エフェクター)分泌複合体。
(注4) ジアシルグリセロール:グリセリンに 2つの脂肪酸がエステル結合を介して結合した分子。宿主細胞膜に局在してセカンドメッセンジャーとして作用する。
(注5) 自然免疫:私達が生まれた時から備えている免疫系。微生物をはじめとする病原体の生体内侵入を認識し排除する生体固有の防御システム。
(注6) 炎症性サイトカイン、ケモカイン:病原体の侵入や自己の異物を認識してその信号を周囲の細胞や組織に知らせ(炎症を起こす)免疫系を活性化させる役割をもつ低分子のタンパク質。それらのタンパク質により炎症部位に好中球など血球系細胞を呼び寄せる。
(注7) プロテオミクス解析:タンパク質の発現動態を網羅的に定量的に解析する、生化学的およびインフォマティクス的な解析手法。本研究では、赤痢菌III型分泌装置によって分泌される未知タンパク質を網羅的に解析し、その中からOspIを同定することができた。
(注8) マイクロアレイ解析:細胞内の遺伝子発現動態を網羅的かつ定量的に測定する解析手法。DNAアレイはその一例。