PRESS RELEASES

記憶の脳回路痕跡 マウスでついに発見!研究成果

記憶の脳回路痕跡 マウスでついに発見!
|
平成26年3月17日
東京大学大学院薬学系研究科
1.発表者:
池谷裕二(東京大学大学院薬学系研究科 薬学専攻 准教授)
2.発表のポイント:
◆ニューロン(神経細胞)で抑制性シグナルに打ち勝つほどの大きな興奮性シグナルが受け取られることにより記憶は思い出される(再生される)ことが分かりました。
◆学習後のマウスの脳スライス標本を調べることで、記憶の痕跡がニューロン間の信号伝達の増強により脳回路に保存されていることを発見しました。
◆記憶が脳内でどのようにしてできるのかを理解することで、逆に、記憶ができない疾患ではどのような問題が生じているのかを解明でき、認知症治療の糸口を探ることができます。
3.発表概要:
東京大学 大学院薬学系研究科の池谷裕二准教授らの研究グループは、脳回路の中の記憶にかかわるニューロンで、興奮性シグナルが増強することが、記憶痕跡の実態であることを証明しました。
一度つくられた記憶は、その後ノンレム睡眠(注1)時に脳内で再生(リプレイ、注2)されることが知られています。しかし、どのような形で記憶痕跡が脳回路に埋め込まれ、脳がどのようにしてその記憶痕跡を再び取り出し再生するかは、いまだに知られていませんでした。
池谷准教授らは、記憶にかかわったニューロンを、そうでないニューロンとは区別できる特殊な遺伝子改変マウスの標本を用いて、記憶に関わったニューロンが優先的に活動しやすくなることで記憶の再生が起こることを示しました。これまで、脳回路ではニューロンの興奮(アクセル)と抑制(ブレーキ)は広くバランスが取れていることが常識でした。しかし、記憶にかかわったニューロンは抑制性シグナルに打ち勝つほどの大きな興奮性シグナルを受け取ることで、記憶を再生させることが明らかになりました。さらに記憶は「LTP(注3)」によって脳回路に保存されていることも証明しました。
本研究成果により、脳が極めて精細な興奮性調節に基づいて記憶を再生するという画期的な発見がもたらされ、記憶のメカニズムの解明に向けた大きな研究の進展が得られました。これは、脳が記憶を再生する仕組みに関するデカルト以来350年の謎を解決したのみならず(附論参照)、今後、認知症など記憶ができない疾患ではどのような問題が生じているのかを解明する手がかりとなることが期待されます。
本研究成果は、2014年3月16日(米国時間)発行の米国科学誌「ネイチャー・ニューロサイエンス」オンライン版に掲載されます。
4.発表内容:
<研究の背景と経緯>
脳は神経細胞(ニューロン)の集まりです。しかし、単にニューロンを集めれば脳が機能するわけではありません。どう集まるか、つまり、どう回路を作るかが重要です。「脳回路」に着目した研究は、必要であることは以前より認知されていたものの、過去ほとんど行われていませんでした。それは、これまで適切な実験手法がなかったためです。東京大学 大学院薬学系研究科の池谷裕二准教授らは「多ニューロン画像法(注4)」を開発し、改良することで、どのような形で記憶痕跡が脳回路に埋め込まれ、脳がどのようにしてその記憶痕跡を再び取り出し再生するかという難題に挑みました。
「夢見る」脳スライス標本の発見
今回の研究成果に至った秘訣は「夢見る脳スライス」の発見にあります。記憶するときに活動したニューロンを、人工的に活性化させると強制的に記憶が思い出されることが、2年前に利根川進らの研究によって明らかにされています(Nature 484:381-385, 2012)。この事実は、記憶に関わったニューロンそのものが「記憶痕跡」であることを意味しています。一方、現実の脳は、この記憶痕跡を(人工的な刺激でなく)自然に再生(リプレイ)します。そして、近年の研究により記憶痕跡の再生は「鋭波(注5)」に現れていると考えられています。
「鋭波」はノンレム睡眠中によく観察される脳波の一種で、記憶や学習を司る脳部位である海馬においても観察されることが古くから知られています。また「鋭波」は、一度つくられた記憶を脳回路に定着させるために重要な働きを担っていると考えられています。興味深いことに、「鋭波」が発生している時には、一度つくられた記憶が脳内でリプレイされていることが近年の研究で明らかになってきました。しかしながら、脳にどのような形で記憶痕跡が残り、どのような仕組みで一度たくわえた記憶痕跡を再び取り出し、記憶をリプレイすることができるのかについては明らかではありませんでした。
記憶をリプレイする仕組みを明らかにするためには、記憶にかかわったニューロンと、そうでない大多数のニューロンとを区別する必要があります。従来のような固定された脳標本を観察する手法では、記憶のリプレイのような“生きたままの動物の脳”で起こる現象について明らかにすることは不可能でした。そこで今回、記憶にかかわったニューロンを蛍光タンパク質で生きたまま標識し、その後にマウスから海馬を取り出し、スライス標本にしたところ、このスライス標本は「鋭波」を自然に生じていることがわかりました。つまり、取り出された海馬は、自然と「睡眠中」の状態になるのです。
さらに重要なことに、眠っている海馬のスライス標本が「鋭波」を発生している最中に、記憶に関わったニューロンが活性化されていたのです。つまり、かつてマウスの頭の中にあった頃の記憶を、海馬のスライス標本になっても、リプレイさせる(≒夢見る)ことができるというわけです。この「夢見る脳スライス」の発見を弾みとして、「では一体、何がこの記憶のリプレイを引き起こしているのか」「記憶に関わるニューロンと関わっていないニューロンとでは何が異なるのか」という重要な問いを追求できるようになり、本研究の布石となりました。
<研究方法と発見の内容>
記憶にかかわったニューロンを標識できるArc-dVenusマウス(注6)からスライス標本を作製し、「鋭波」が発生しているときの、ニューロンの活動を観察しました。記憶にかかわったニューロンとそうでないニューロンを区別しながら活動を観察するために、独自に開発した特殊なカルシウム蛍光指示薬「CaSiR-1」を用いて、カルシウムイメージングを行いました。すると、記憶にかかわったニューロンのほうが、そうでないニューロンに比べて、「鋭波」が発生しているときに活動しやすくなっていることが判明しました。これは、「鋭波」が発生しているときには、まさに記憶のリプレイが起こっていることを意味しています。この記憶のリプレイ現象はLTPの維持過程を阻害することで観察されなくなりました(注7)。以上の結果から記憶は、LTPによって痕跡として脳回路に保存されることで、リプレイされることが示唆されました。
この記憶のリプレイがどうして起こるのかをさらに詳細に調べるために、ひとつひとつのニューロンからパッチクランプ記録(注8)を行い、ニューロンに入ってくる興奮性シグナル、および抑制性シグナルをそれぞれ詳細に観察しました。この観察により、記憶のリプレイに関係するニューロンでは、興奮性シグナルと抑制性シグナルのバランスが興奮性側にシフトしていることが判明しました。さらに、ニューロンに人工的なシグナルを注入し、「鋭波」が起こっている際の神経活動のシミュレーションを行うと、興奮・抑制のバランスが興奮性側にシフトしているシグナルを受けることこそが、ニューロンの活動を再現良く引き起こすことができることも明らかになりました。
そこで、興奮・抑制のバランスがどのような仕組みでつくられるのかについて、さらに詳細に調べたところ、ひとつひとつのニューロンはバラエティ豊かな、個性的な興奮性シグナルを受け取る一方で、それぞれのニューロンが受け取る抑制性シグナルは均一であり、脳回路内の興奮性シグナルの平均レベルを反映することが判明しました。この均一な抑制性シグナルの由来として、抑制性介在ニューロンの一種であるパルバルブミン陽性インターニューロン(注9)に着目し、その活動を観察したところ、パルバルブミン陽性インターニューロンは、脳回路内の興奮性シグナルを網羅的に受け取り、興奮性レベルを検知してそれに合わせた出力を行うことで、まわりのニューロンに均一な抑制性シグナルを送っていることが示唆されました。
今回の実験結果から、以下のような流れで記憶が再生されると示唆されます(図1も参照ください)。記憶がつくられる時に活動したニューロンではLTPが起こり、上流のニューロン集団との結合性が高まります。そのため記憶にかかわったニューロンは平均よりも大きな興奮性入力を受けます。次に、パルバルブミン陽性インターニューロンは脳回路全体に入ってくる興奮性シグナルを網羅的に受け取ることで、脳回路全体の興奮性レベルを検知します。そのレベルに応じた発火活動を行うことで、近傍にあるニューロンに一様に抑制シグナルを送ります。このような多様性に富んだ興奮性シグナルと、一様な抑制性シグナルの組み合わせにより、記憶にかかわった一部のニューロンのみが活動することができ、記憶がリプレイされるという仕組みです。つまり記憶痕跡とは「抑制に打ち勝つことのできるほどの興奮をうみだすシナプス増強」であることが示唆されます。
<今後の展開>
本研究では、興奮性シグナルが抑制性シグナルに打ち勝つことができるようになることこそが記憶のリプレイを支える仕組みであることが明らかになりました。この発見は、脳回路は記憶をリプレイするために、興奮・抑制のバランスを適切に調節できるようにきわめて巧妙に配線されていることを示しています。脳回路の興奮・抑制バランスの精密な調節は記憶・学習能に関わる基本的な生理メカニズムであると示唆されます。
記憶の痕跡はLTPとして脳回路に保存されている一方で、刻まれた記憶が正確にリプレイされるためには抑制性ニューロンによる脳回路の興奮性シグナルの調節が重要であると示唆されます。これは、従来はあまり注目されてこなかった抑制性ニューロンが記憶障害の新たな治療の標的となりうる可能性を高めるものです。今後は、認知症、統合失調症、うつ病など、記憶の変調を伴う疾患において、興奮・抑制のバランスがどのように変化しているかを観察していく新たなアプローチが、精神神経疾患の病態に有益な解釈をもたらすと期待されます。
5.発表雑誌:
雑誌名:Nature Neuroscience(ネイチャー・ニューロサイエンス)
論文タイトル:Unbalanced excitability underlies offline reactivation of behaviorally activated neurons (アンバランスな興奮性が記憶痕跡をかたちづくる)
著者:Mika Mizunuma, Hiroaki Norimoto, Kentaro Tao, Takahiro Egawa, Kenjiro Hanaoka, Tetsuya Sakaguchi, Hiroyuki Hioki, Takeshi Kaneko, Shun Yamaguchi, Tetsuo Nagano, Norio Matsuki, Yuji Ikegaya*
6.問い合わせ先:
池谷 裕二(イケガヤ ユウジ)
東京大学 大学院薬学系研究科 薬学専攻 薬品作用学教室 准教授
7.用語解説:
(注1)ノンレム睡眠:睡眠には主に2つの状態が交互に現れることが知られており、そのうちの一つ。急速な眼球運動や覚醒時と同じような脳波を示すレム睡眠ではない、睡眠状態を指します。
(注2)リプレイ:
記憶学習時の神経細胞集団の発火活動パターンが、後に圧縮再生される現象。
(注3)LTP(Long-term potentiation):長期にわたり、ニューロンからニューロンへ信号が伝達しやすくなる現象を指します。ニューロン間の接合部位(シナプス)が示す可塑性の一種であり、記憶の素過程と考えられています。長期増強とも呼びます。
(注4)多ニューロン画像法:従来の電気生理学的な手法とは異なり、カルシウム蛍光色素を用いることで、同時に多数のニューロンから発火活動を記録する方法。ここでは200個以上のニューロンから一斉に記録を行っています。
(注5)鋭波:
ノンレム睡眠時や静止覚醒時に海馬でよく観察される脳波。周波数が5-30Hzほどの鋭波には、約200Hzのリップル波が重なってあらわれます。鋭波の発生時に記憶がリプレイされることは記憶の固定化に重要な役割を果たしていると考えられています。
(注6)Arc-dVenusマウス:
Arcプロモーターの下流にdVenus配列を配置した遺伝子改変マウス。Arcは最初期遺伝子の一種で、神経活動依存的に発現する。dVenusは改変型GFPであるVenusに不安定領域を付加し、分解性を高めたもの。このマウスでは、活動したニューロンがその後数時間にわたってdVenusを発現します。
(注7)この記憶のリプレイ現象はLTPの維持過程を阻害することで観察されなくなりました:
本研究では、LTPの維持過程を阻害するペプチドZIP(zeta inhibitory peptide)を用いました。ZIPは一度起こったLTPを元に戻してしまう働きを持つことが知られています。
(注8)パッチクランプ記録:
先端に細孔の空いたガラス製の微細電極の先端部分に、特定の細胞の細胞膜を顕微鏡下で実験的に貼り付け電位差や伝導性などの電気的特性を調べる手法。
(注9)パルバルブミン陽性インターニューロン:
カルシウム結合タンパク質パルバルブミンを発現する抑制性介在ニューロン。鋭波発生時に活動率が上昇することが既に知られています。抑制性介在ニューロンは抑制性伝達物質GABAを放出して、興奮性ニューロンに抑制性シグナルを送ります。
8.添付資料:
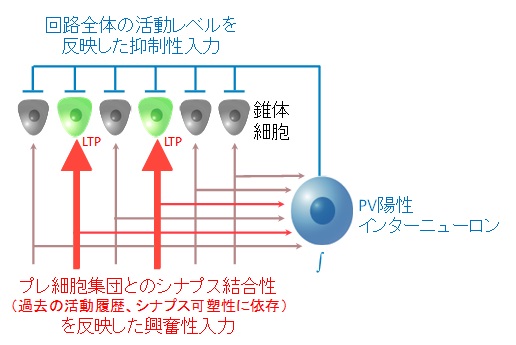
図1:記憶のリプレイの仕組み
記憶にかかわったニューロン(緑)は、LTPにより平均よりも大きな興奮性シグナルを受け取る。そのため、回路の活動レベルを検知したパルバルブミン(PV)陽性インターニューロンから送られる抑制性シグナルに打ち勝つことができる。またLTPによる興奮性シグナルの増強こそが記憶の痕跡をかたちづくる。
<本研究の主な助成事業>
最先端・次世代研究開発支援プログラム
革新的技術を用いて脳疾患を理解する「システム薬理学」の創成
研究代表:池谷裕二(東京大学大学院薬学系研究科)
科学研究費補助金
新学術領域『こころの時間学 ―現在・過去・未来の起源を求めて』
研究代表:北澤茂(大阪大学大学院生命機能研究科)
(池谷は、計画研究者として参加)
科学研究費補助金
新学術領域『メゾスコピック神経回路から探る脳の情報処理基盤』
研究代表:能瀬聡直(東京大学大学院新領域創成科学研究科)
(池谷は、連携研究者として参加)
脳科学研究戦略推進プログラム
精神疾患バイオマーカーの開発とDecNef等による臨床応用のための技術基盤整備
研究総括:川人 光男(国際電気通信基礎技術研究所脳情報通信総合研究所)
(池谷は、共同研究者として参加)
<本研究の共同研究機関>
本研究では京都大学 医学部、岐阜大学 医学部から供与された実験材料を一部に用いています。実験施行・データ解析、論文執筆は東京大学 大学院薬学系研究科で行いました。
附論:なぜデカルト以来なのか
デカルトは近代的な思弁アプローチにもとづいて脳の動作原理を正確に射抜いた最初の哲学者です。1649年に著した『情念論』でデカルトは、記憶のメカニズムについて考察し、次のように記しています。
心が或る事柄を想起しようと欲した場合、(中略)、思い出そうとする対象が残した痕跡の存在する箇所に出会うまで、脳の各所に精気を押し流すのである。けだしこの痕跡とはかつて問題の対象が現れたために精気がそこから流れ出した脳気孔にほかならず、その結果、この痕跡は精気が到達した場合、ふたたび同様にして開くことが、他の気孔と比べてはるかにたやすくなっているのである。したがってこの気孔に出会った精気は、(中略)、この対象こそ心の思い出そうとしていた物であることを心に教えるのである。
当時、シナプスはおろか神経細胞すら発見されていなかったため、このような難解な文章になったと思われます。そこで、「精気」を「活動電位」に、「気孔」を「シナプス」に読みかえると、以下のようになります。
ある事柄を思いだそうとしたとき、脳にたくえられた過去の記憶(痕跡)を探すために、脳の各所に活動電位を送りこむ。痕跡とは、過去に活動電位が通過したシナプスのことで、その結果として、次に活動電位が到達したとき、このシナプスが活動することが、はるかにたやすくなっている。したがって、このシナプスにたくわえられた記憶こそが、いままさに思いだそうとしていた物として想起されるのだ。
太字で示したデカルトの推察こそ、今回の発見した現象そのものです。記憶については、古代ギリシャ以来(アリストテレスなど)、多くの識者が洞察を重ねてきましたが、350年前のデカルトほど本質をついた洞察を示した人はいません。